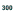Научные статьи
К разработке методик породоиспытаний и порядка госрегистрации, генетической паспортиз
Назревшая необходимость экологически обоснованной и экономически рентабельной биоконверсии органических отходов переработки биологической продукции вызвала распространение технологии вермикомпостирования и в России. Основным объектом вермикультивирования на первых порах стал Красный Калифорнийский гибрид(ККГ), завезенный в Россию через Ивано-Франковск (Украина) из Польши и Венгрии в 1990- 91 г., относимый к бесполосой форме компостного (навозного) червя, описанной как подвид Eisenia foetida andrei Bouche,1972. Полосатая форма Eisenia foetida foetida Savigny 1826, для которой не найдено отличий от него по морфологическим признакам (Bouche,1972; Jaenike, 1982), давала с ним нефертильное гибридное потомство, отличаясь и по составу аллелей пяти локусов изоферментов, что характерно для самостоятельных видов (Jaenike, 1982).

Наряду с ККГ в целом ряде районов России начали использовать для компостирования и местных навозных червей (Морев, 1982: Андреев А.Г., Кодолова О. П., Нефёдов Г. Н., Трувеллер К. А., Серкутан К. Н.1992; Игонин, Титов, 2002), разнообразных по продукционным характеристикам (Кодолова и др., 1993) и подразделяемых по частотам аллелей 4 полиморфных локусов аллозимов на «южную» и «северную» географические группировки (Болотецкий и др., 1994) неясного таксономического статута (Болотецкий, Кодолова, 2002).
К настоящему времени в России появился ряд промышленных линий, полученных на основе местных популяций гибридизацией и селекцией на разные кормовые субстраты (Кодолова О.П., Болотецкий Н.М., Правдухина О.Ю., Андреев А.Г.1994а,б; Трувеллер К.А.1996; Шония А.М.1996; Болотецкий Н.М., Батурин А.Л., Жуковская Е.А., Кодолова О.П., Косевич И.А., Нефедов Г.Н., Правдухина О.Ю., Путилова Э.В., Трувеллер К.А1997; Игонин А.М., Титов И.Н.2002; Мамеева В.Е., Просянников Е.В., Трувеллер К.А., 2002;).
Однако до государственной регистрации и описания селекционных достижений для нового объекта культивирования, каким является компостный (навозный) червь и, возможно, ряд других видов дождевых червей, необходимо разработать и утвердить соответствующие специфике биологии и использованию объекта методики оценки отличимости, однородности и стабильности пород и показатели продуктивности, соответствующие данному виду (Эрнст и др.,1987), а также формы их анкетирования согласно общим утвержденным «Правилам составления и подачи заявки на допуск селекционного достижения к использованию» (от 14.10.1994 №2-01/4) и разъяснению к ним (10.04.1995 №12-04/4).
Предлагаются для обсуждения следующие показатели и методики при описании и заявке в реестр пород компостных червей Eisenia foetida.
Диагноз вида Eisenia foetida (Savigny 1826) (Oligochaeta, Annelida, Lumbricidae) заявляемой породы следует уточнить по определителю Перель, 1979 и свериться с его описанием, поскольку, например, только род Eisenia в фауне СССР представлен 20 видами. Русское название вида Eisenia foetida (Savigny,1826) - навозный червь (Чекановская,1960);. Практически важным представляется описание окраски тела – как отличительного признака подвидов Eisenia foetida andrei, Bouche,1972-бесполосого и Eisenia foetida foetida (Savigny, 1826) - полосатого (Bouche,1972; Jaenike, 1982), и его подтверждение по биохимико-генетическим маркерам (БХГМ) (Lap-2, Pgm, Pgi, Est-1, Est-3, (Jaenike, 1982)), а также географических форм «Северные» и «Южные» по БХГМ (Odh, Lap, Cat, Pgm ( Болотецкий, 2004; ), а также по Est-3, Pr-3,5 и Mdh).
Скрещиваемость, ввиду отсутствия четких видовых диагностических морфологических признаков рода Eisenia (Перель, 1979) и противоречивости сведений о таксономическом статуте foetida и andrei (Jaenike, 1982), необходимо включить в испытание пород и проводить до третьего поколения включительно согласно методике определения плодовитости в попарных скрещиваниях в стаканах на последовательных потомствах не менее чем на 30 парах червей в каждом из 4 поколений породы и включая для гибридных кроссов и форм основателей.
Отличимость пород червей возможна пока только по перспективным БХГ маркерам Odh, Lap, Cat, Pgm, (Болотецкий Кодолова, 2002; ) Lap, Pgm, Est-3, Mdh, Ldh-1,2, Glcdh, Gpdh-2 (Jaenike, 1982; Robotti,1982; Diehl,1988; McElroy et al., 1995) и эколого-продукционным показателям (ЭПП).
Описание стандартных методик БХГМ: Диск-электрофорез в вертикальных блоках ПААГ (Маурер, 1971; Трувеллер, Нефедов, 1974). Проявление и номенклатура БХГМ (Корочкин и др., 1977; Болотецкий, Кодолова, 2002; Болотецкий, 2004; Jaenike, 1982; Diehl,1988; Richardson et al., 1986)
Однородность по БХГМ: равновесие по соотношению генотипов наблюдаемых ожидаемым из частот аллелей трех-четырех локусов аллозимов. Достоверная (P>0,99) неоднородность по соотношению аллелей и генотипов трех-четырех локусов аллозимов при сравнении с другими породами и/или популяциями дождевых червей.
Стабильность по БХГМ: равновесие ХВ и однородность по аллелям и генотипам БХГМ всех отводок и поколений породы, как было показано, например, для трех разных отводок ККЧ (Болотецкий, Кодолова, 2002; Болотецкий, 2004).
Описание стандартных методик получения эколого-продукционных показателей.
Представляют следующие продукционные характеристики заявляемых и сравниваемых пород и исходных популяций червей:
· число коконов, полученных от каждого экземпляра червя за 1 неделю (кок./нед.);
· число личинок, вылупившихся из одного кокона (лич./кок.);
· время инкубации коконов с точностью до 3-4 суток, (сут.).
Оценочный показатель плодовитости рассчитывают по исходным данным как среднее значение числа личинок от зрелого экземпляра за неделю и в сутки:
· коллумелярный вес - средняя масса оособи группы зрелых минимальных (г/экз.);
· срок наступления половой зрелости (сут.) от откладки кокона и от вылупления личинки до появления пояска - клителлума.
Возрастную структуру популяций наблюдают в течение 2-3 циклов развития до созревания или 41 недели при комнатной температуре на стандартном субстрате в ящичных культурах (смешанной пробой до дна из 3 ящиков), либо на ложах, грядах (пробоотборником 1 дм2 глубиной 40см смешением трех проб) еженедельно учитывая количество взрослых особей, молодых червей и коконов по нижеприведенным группам:
· Коконы - (яйцевые капсулы) живые и свежие, имеют светло-зелёный цвет, зрелые с личинками коричневого цвета. Отдельно учитывают также пустые и погибшие бурые коконы;
· личинки (Larvae) – вылупившиеся за неделю черви минимального размера со слабой пигментацией кутикулы, но с видимым кровеносным сосудом;
· молодь (Juvinis) –вполне пигментированные черви, превосходящие по размерам личинок, но короче наименьших зрелых особей (min Аd );
· подрост (Vergin) – равные по размеру наименьшим зрелым особям, но лишённые пояска - клителлюма и следа перетяжки от него;
· зрелые минимальные (min Аdultus) – наименьшие черви с пояском или четкой перетяжкой;
· зрелые средние (mid Ad} – черви с пояском на 20% длиннее min Аd и короче max Аd
· зрелые максимальные (max Ad) – черви наибольшей длины ;
Зрелых делят на три размеренные группы условно в пределах каждой выборки, учитывая их численность, вычисляют соотношение этих групп в %% от суммы зрелых и от всех возрастных групп червей, общий вес по гуппам для получения средней массы тела в динамике на протяжение трех циклов развития
Плодовитость и продуктивность определяют на основе числа коконов от экз. за неделю, числа личинок из кокона, среднего времени инкубации кокона , времени развития до созревания и инд. массы тела при созревании при стандартных условиях для испытываемой и двух контрольных популяций.
Число коконов от 1 червя за неделю, учитывают при комнатной температуре в отдельных пластмассовых стаканах объёмом 250 или 500 см3, наполненных стандартным субстратом, в каждый из которых помещают по две неполовозрелые особи (из подроста) по 15 или 30 парам каждой из трех пород.
Время инкубации коконов и число личинок, вылупившихся из одного кокона (Барне, 2002) определяют при комнатной температуре (Reinecke, Kriel, 1981). В чашки Петри диаметром 10 см закладывают на увлажнённую фильтровальную бумагу по 5 или 10 коконов одного срока кладки. Выход личинок после первых 10 суток от закладки учитывают через каждые 3 или 5 дней в течение всего срока инкубации.
Однородность и стабильность породы по ЭПП устанавливается в динамике относительно объектов базы сравнения- ККГ и исходных форм. Оценивают интенсивность роста по живой массе тела (мг) минимальных и максимальных зрелых червей и сроку созревания от выхода личинок, производительность как плодовитость: (коконы, личинки/экз/неделя) и скорость развития от откладки кокона до созревания за принятый период или 4 поколения на стандартном субстрате ЭПП при стандартных условиях.
Сравнение заявляемой породы - кросса (например СТАРАТЕЛЬ (NхS)) - с теми же показателями др. пород (ККГ) базы сравнения и исходных форм (Владимирская местная(N} и Чуйская(S)) в условиях региона; требует достоверного превышения показателей для линии кросса (СТАРАТЕЛЬ) над показателями исходных линий (Владимирская и Чуйская популяции) и породы сравнения ККГ.
Полезные технологические качества породы предусматривают Правилами, в том числе, наличие генетических маркеров, облегчающих технологические и селекционные процессы (например, идентификация, отличимость породы, сохранение стабильности генофонда) для дифференцирования биотехнологических (субстрат, тепловой режим) и генетических (инбридинг, генетическое засорение) причин при снижении продукционных показателей вермикультуры. Рассматривается также приспособленность к конкретным технологиям производства и природным условиям, особенности поведения и воспроизводства, жизнеспособность, продолжительность использования ( в отношение червей – период плодовитости и интенсивного роста поколения). Причем, представляемые полезные технологические качества породы должны быть достоверно выше, чем у пород базы сравнения, в частности, ККГ и исходных форм червей.
Определение обязательной численности племенной части породы, обеспечивающей стабильность генофонда как основы показателей продуктивности и отличимости (минимум 300х100=30000 червей). Производственная минимальная численность заявленной породы червей должна обеспечить эффективное заселение 100 м2 рабочих лож – т.е. 2000000 общей численности всех возрастов.
Цветные фото 9х12 или 13х18 с характерными особенностями основного типа экстерьера для пород E. foetida должны отразить наличие или отсутствие полос по средней части тела, пигментацию зрелых особей, а электрофореграммы показать фракции аллозимов с возможностью идентификации их по относительной подвижности (Rf), диагностичных для Ю и С форм и породы по сравнению с др. породами.
Обоснование на использование породы (технические условия на продукцию, заиинтересованность производства), которое может быть предложено представить при проведении экспертизы заявленной породы, также предстоит разработать для компостных червей в соответствии с направлением вермихозяйств по аналогии с другими объектами культивирования.
Литература
1. Андреев А.Г., Кодолова О. П., Нефёдов Г. Н., Трувеллер К. А., Серкутан К. Н. Перспективы развития вермикультуры на основе селекции. // Тезисы докладов: Материалы международной научно-практической конференции. Днепропетровск, 1992, - 20 с.
2. Барне А. Ж. Динамика сброса коконов у компостного червя Eisenia foetida. Материалы I-й международной конференции «Дождевые черви и плодородие почв». – Владимир, 2002 – С. 7 – 8.
3. Болотецкий Н.М. Об электрофоретических уровнях сходства некоторых видов дождевых червей сем. Lumbricidae. Рукопись деп. в ВИНИТИ 18 июля 1985 г., № 5200-85 деп.
4. Болотецкий Н. М. Внутривидовая генетическая и морфологическая изменчивость дождевого червя Eisenia fetida (Sav.) Владимир,2004.
5. Болотецкий Н.М., Батурин А.Л., Жуковская Е.А., Кодолова О.П., Косевич И.А., Нефедов Г.Н., Правдухина О.Ю., Путилова Э.В., Трувеллер К.А.// Популяционные биохимико-генетические основы селекции компостного червя (Eisenia foetida Sav.). Тез.докл. Муждунар.науч.-практич.конф. "Перспективы биологической переработки органических отходов с помощью вермикультивирования". Рязань, 18-21 марта 1997, с.17-20.
6. Болотецкий Н. М., Кодолова О. П., Нефёдов Г. Н., Правдухина О. Ю., Трувеллер К. А. О технологии получения гибридных линий навозного червя Eisenia foetida (Sav.) // Тезисы докладов II Международной конгресса. Биоконверсия органических отходов народного хозяйства и охрана окружающей среды. – Ивано-Франковск V 1992. – С. 17-18.
7. Болотецкий Н.М., Кодолова О.П., Правдухина О.Ю. Генетическое разнообразие навозного червя Eisenia foetida (Sav.). Изв.РАН.Сер.биол. 1994. №3 с.404-410.
8. Зеленкова А.А. Разработка селекционных индексов и совершенствование методов оценки племенной ценности производителей в мясном скотоводстве. Автореф. дис. кан.с.-х. наук: 06.02.01. – Пернановка, 1996 – 22
9. Игонин А.М., Титов И.Н. Переработка органических отходов с помощью новой промышленной линии дождевых (компостных) червей «Владимирский гибрид» (Старатель). Материалы I-й международной конференции «Дождевые черви и плодородие почв». – Владимир, 2002 – С. 7 – 8.
10. Кодолова О.П., Болотецкий Н.М., Правдухина О.Ю., Андреев А.Г. О необходимости селекции навозного червя для вермикультивирования. Материалы международного симпозиума, 1994. – С. 29 – 30.
11. Кодолова О.П., Болотецкий Н.М., Правдухина О.Ю., Андреев А.Г. О результатах селекции навозного червя Eisenia foetida (Sav.), Материалы международного симпозиума 1994. – С. 29 - 30.
12. Кодолова О.П., Стриганова Б.Р., Сидорова Т.И. Сравнительное исследование продуктивного потенциала локальных поселений компостного червя Eisenia foetida (Savigny 1826) (Oligochaeta, Lumbricidae) Изв.РАН.Сер.биол.1993 №4 с.858-867.
13. Корочкин и др. Генетика изоферментов. М.: Наука, 1977, 280 стр.
14. Мамеева В.Е., Просянников Е.В., Трувеллер К.А. О получении Брянской линии дождевых компостных червей для вермикультивирования. Научно-практическая конференция: «Использование достижений современной биологической науки при разработке технологий в агрономии, зоотехнии и ветеринарии». Брянск. 2002.
15. Матвеева В.Г., Гвоздева О.А, Коновалова М.П., Фирова С.А. О размножении и развитии дождевых червей Eisenia foetida Sav., Nacopeilus caliginosus Sav. F. fipyca в лабораторных условиях. //Структура и динамика популяций почв наземных беспозвоночных животных, 1990 – С. 104-108.
16. Маурер Г. Диск-электорофорез. М.: Мир, 1971, 242 стр.
17. Морев Ю.Б. Разведение дождевых червей на отходах животноводства. // Сер. Химико – технологические науки. / Фрунзе: Изд. АН Кирг. ССР. 1989. № 1. - С. 60 – 64.
18. Перель Т.С. Распространение и закономерности распределения дождевых червей. – М.: Наука, 1979. – 70 с.
19. Просянников Е.В., Трувеллер К.А., Мамеева В.Е. и др. Вермитехнология и селекция компостных червей на Брянщине. Материалы I-й международной конференции «Дождевые черви и плодородие почв». – Владимир, 2002 – С. 21 – 24.
20. Светлов П.Г. 1928. Исследования над развитием дождевых червей. Тр. Особ. зоолог. лабор. и Севаст. ст. АН СССР, сер. 2, № 11 – 13 : С. 95 – 329.
21. Трувеллер К.А. Выведение региональных промышленных линий вермикультур на основе селекционно-генетической коллекции навозного червя. // Тезисы докладов Всероссийской научно-практической конференции «Перспективы биологической переработки органических отходов с помощью вермикультуры. Опыт обучения специалистов-вермитехнологов». Рязань, 1996, - С. 9-11.
22. Трувеллер К.А., Нефедов Г.Н. // Многоцелевой прибор для вертикального электрофореза в параллельных пластинах полиакриламидного геля. // Науч. докл. высш. школы, сер. биол. науки,1974, N9,137-140.
23. Трувеллер К.А., Нефёдов Г.Н. Ускоренное получение гибридных местных популяций компостных червей Eisenia foetida (Lumbricidae, Oligohaeta) для экоциклинга органических отходов. Материалы I-й международной конференции «Дождевые черви и плодородие почв». – Владимир, 2002 – С. 7 – 8.
24. Чекановская О.В. Дождевые черви и почвообразование. М.; Л.: Изд-во АН СССР, 1960. 206 с.
25. Шония А.М. Генетические центры вермикультивирования. Современные достижения биотехнологии. Ставрополь, 1996, - С. 17 – 18.
26. Шурова Н.М. Видовое разнообразие олигохет и чистота культуры Eisenia foetida. Материалы международного симпозиума 1994. – С. 30-31.
27. Эрнст Л.К., Кравченко Н.А., Солдатов А.П. И др.; Под ред. Кравченко Н.А. // Племенное дело в животноводстве. – М.: Агропромиздат, 1987. – 287 с.: ил.
28. Bouche M.B. Lombriciens de France: Ecologie et systematique. Paris, 1972, pp. 380-382.
29. Diehl W. J. Genetics of carbohydrate metabolism and grouth in Eisenia foetida (Oligochaeta: Lumbricidae). Heredity 61(1988) 379-387
30. Grove A.J. and L.F. Cowley. 1926. On the reproductive processes of the brandling worm Eisenia foetida. Quart. Journ. Micr. Sci., 70 : 559 – 581.
31. Jaenike J. “Eisenia foetida” is two biological species. Megalodrilogica 4(1982) 6-8.
32. McElroy T.C., Presley M.L., Diehl W.J. Genotypes at seven polymorphic allozymes loci interact with the environment to affect growth rate in juvenile earthworms (Eisenia foetida). Abstr. Annu. Meet. Amer. Soc. Zool., Chicago, Dec., 26-30, 1995.
33. Reinecke A.J., Kriel J.R. The influens of constant and diurnally fluctuating temperatures on the cocoon prodaction, hatching time, and number of hatchings of Eisenia foetida (Lumbricidae, Oligochaeta) // Workshop on the role of Earthworms in the Stabilization of Organic Residues. Kalamazoo, Michigan, 1981. - Vol. 1. P. 166.
34. Robotti C.A. Biochemical polymorphism of earthworms. 4. Enzymes of Eisenia foetida Andrei (Bouche )(Annelida, Oligochaeta) . Monit. Zool. Ital. 1982 №16 з.341-344
Источник: Материалы II научно-практической конференции "Дождевые черви и плодородие почв"
|